
-

生物通官微
陪你抓住生命科技
跳动的脉搏
生物通官微
陪你抓住生命科技
跳动的脉搏
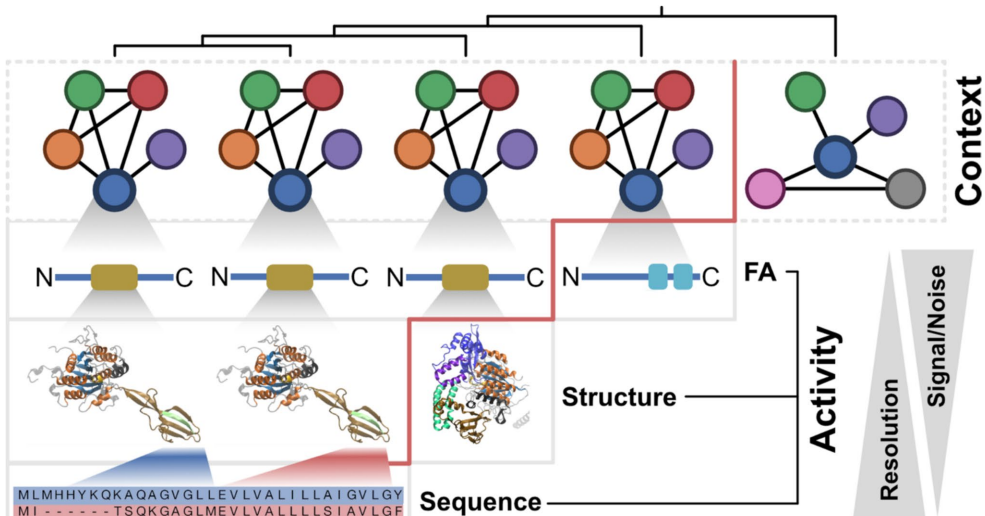
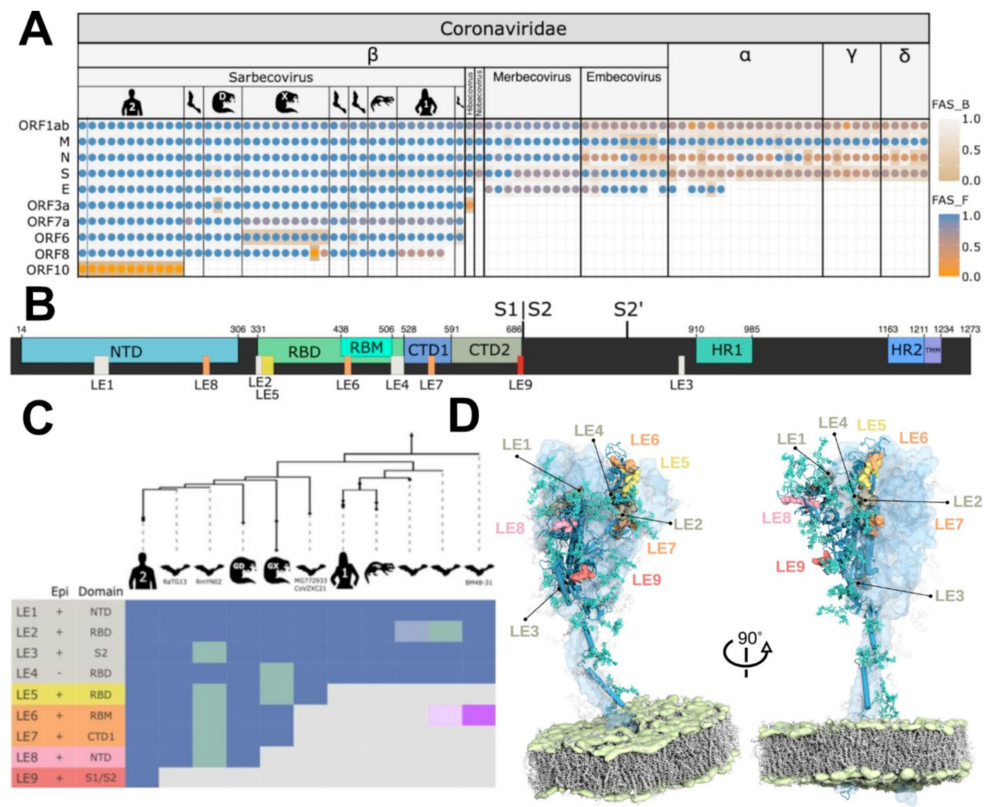
综述:直系同源物功能演化的多层级视角
《Journal of Molecular Evolution》:A Multi-level Perspective on the Evolution of Orthologs and Their Functions
【字体: 大 中 小 】 时间:2025年10月19日 来源:Journal of Molecular Evolution 1.8
编辑推荐:
这篇综述挑战了直系同源物(orthologs)必然功能等同的传统观点,主张将其视为需严格检验的零假设。作者提出一个多层级分析框架,整合功能背景(如代谢通路)和生化活性(如蛋白结构域)的证据,以系统评估功能分化。该文为大规模比较基因组学中精准进行功能注释转移(Gene Ontology, KEGG)提供了方法论指导,并强调开发量化功能分化的新指标(如FAS评分、AlphaMissense)是未来重要方向。
 生物通微信公众号
生物通微信公众号
知名企业招聘

