SUPERMAN通过EAR基序招募共抑制子TOPLESS调控花器官边界的分子机制
《BMC Plant Biology》:SUPERMAN requires its EAR motif to recruit the corepressor TOPLESS and to function as a cadastral regulator in floral patterning
【字体:
大
中
小
】
时间:2025年11月09日
来源:BMC Plant Biology 4.8
编辑推荐:
本研究针对植物花器官发育中边界基因表达调控的机制问题,聚焦转录抑制因子SUPERMAN(SUP)的EAR基序功能。研究人员通过遗传学、生物化学和基因组编辑技术,首次证实SUP通过其EAR基序直接招募共抑制子TOPLESS(TPL),共同抑制B类花器官基因APETALA3(AP3)/PISTILLATA(PI)的中央扩散,从而维持花轮3-4边界。该发现揭示了植物发育中转录抑制复合物的精确调控机制,为理解植物器官边界形成提供了新范式。
在植物发育的精密调控网络中,花朵如何精确划定不同器官的边界一直是个迷人的谜题。拟南芥的花朵由四轮同心圆排列的器官组成:最外层是萼片,向内依次为花瓣、雄蕊和心皮。经典的ABC模型告诉我们,这三类花器官身份基因的组合表达决定了器官的命运。但问题在于――这些基因的表达边界如何被严格限定?一旦边界失控,就会导致器官身份错乱,比如本该长心皮的位置长出额外的雄蕊。
这就是SUPERMAN(SUP)蛋白大显身手的舞台。早在1990年代,科学家就发现SUP突变体会产生"超级雄蕊"――花朵中心会额外长出多轮雄蕊,同时心皮身份受损。研究表明,SUP作为转录抑制因子,其羧基末端含有一个关键的EAR基序(ERF-associated amphiphilic repression motif),但这一基序在体内如何发挥作用,以及与哪些分子伙伴协作,始终是未解之谜。
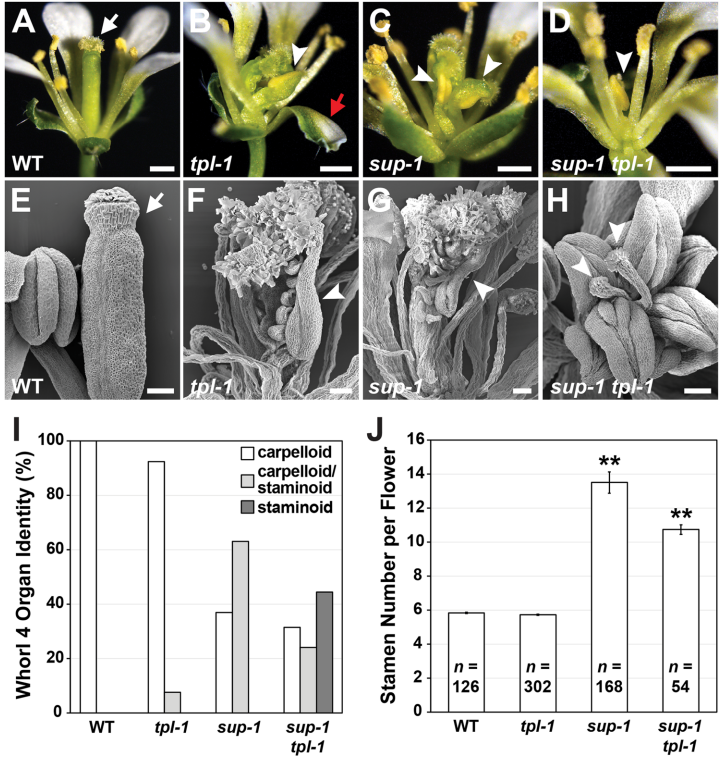
Wokasch等人在《BMC Plant Biology》发表的研究正是针对这一科学问题展开。他们发现,原本被认为主要在外轮起作用的共抑制子TOPLESS(TPL),其实在花朵中心也扮演着关键角色。tpl-1突变体的心皮中出现了雄蕊组织,这种雄性生殖命运向中心扩散的现象与sup-1突变体惊人相似。更令人惊讶的是,当两者同时突变时,花朵中心几乎完全丧失了心皮特征,显示出强烈的协同效应。
那么,SUP和TPL是否真的在分子层面直接对话?研究人员通过酵母双杂交实验给出了肯定答案。SUP与TPL之间存在强烈的物理相互作用,而这一相互作用完全依赖于SUP的EAR基序。当研究人员将EAR基序中的关键亮氨酸残基突变为丙氨酸时,SUP就失去了与TPL结合的能力。
为了验证EAR基序在活体中的重要性,研究团队运用CRISPR-Cas9基因组编辑技术,精准靶向了SUP基因的EAR基序编码区。他们获得了两个新颖的等位基因:sup(DLELL)仅缺失3个碱基,导致EAR基序从DLELRL变为DLELL;sup(△EAR)则缺失42个碱基,完全消除了EAR基序。令人震惊的是,即使是单个氨基酸的细微改变,也足以导致强烈的花表型缺陷。
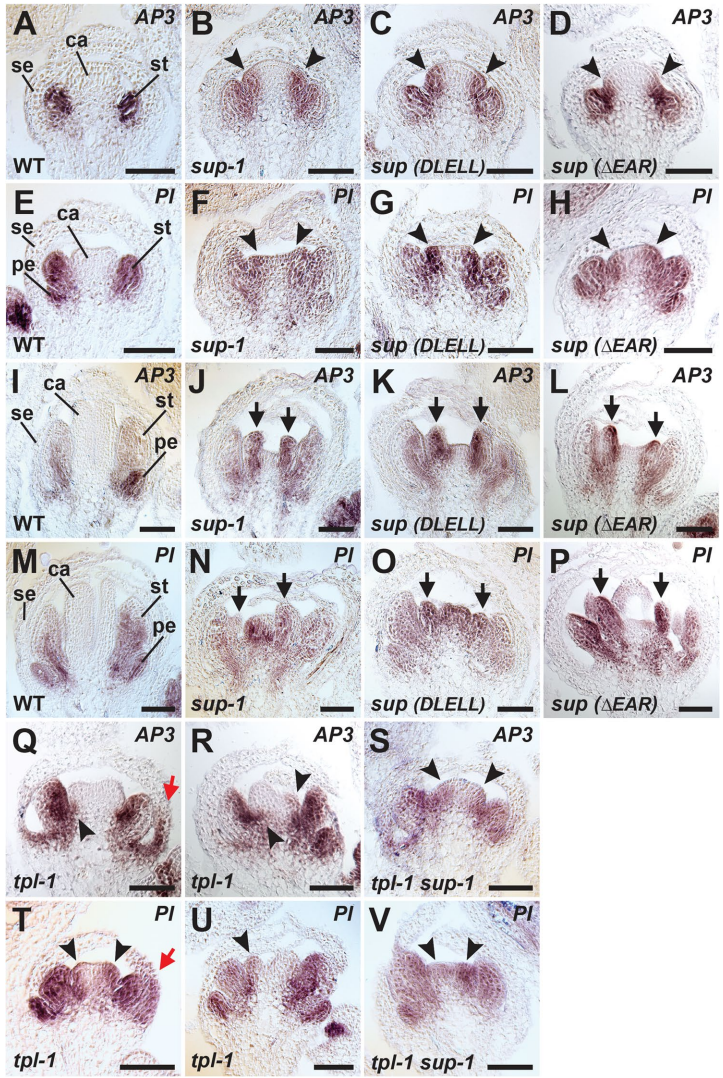
这些基因组编辑的突变体表现出与强等位基因sup-1几乎相同的缺陷:雄蕊数量增加,心皮中出现雄蕊组织。通过RNA原位杂交,研究人员进一步证实,sup(DLELL)和sup(△EAR)花朵中B类基因AP3和PI的表达确实从第3轮扩散到了第4轮,这与sup-1的表型完全一致。
最关键的是,这些由CRISPR产生的SUP变异体蛋白在体外pull-down实验中显示,它们与TPL的结合能力严重受损。即使是细微的sup(DLELL)突变,也将TPL结合能力降低至野生型的32%。这完美解释了为什么这些突变体会出现强烈的发育缺陷。
本研究主要采用了以下关键技术方法:利用CRISPR-Cas9基因组编辑技术构建EAR基序特异性突变的拟南芥株系;通过酵母双杂交和体外pull-down实验验证蛋白质相互作用;采用RNA原位杂交和RT-qPCR分析基因表达模式;使用扫描电镜观察花器官形态;并通过对tpl、sup单双突变体的遗传学分析揭示功能协同效应。
研究发现tpl-1突变体与sup-1类似,在花轮4器官中出现雄蕊组织嵌合体,表明TPL与SUP共同抑制中心轮雄性命运。双突变体表型增强显示协同作用。
酵母双杂交和体外pull-down实验证实SUP与TPL直接结合,且该结合依赖于EAR基序的完整性。亮氨酸残基的突变显著削弱相互作用。
CRISPR编辑的sup(DLELL)和sup(△EAR)株系表现出与sup-1相似的花器官缺陷和B类基因表达扩散,证明EAR基序对SUP功能不可或缺。
RNA原位杂交显示所有sup等位基因和tpl-1背景中AP3/PI表达均突破花轮3-4边界,支持SUP-TPL复合物共同维持表达边界的模型。
分子实验证实CRISPR产生的SUP变异体蛋白与TPL结合能力受损,为表型缺陷提供了机制解释。
这项研究最终构建了一个完整的调控模型:SUP作为花器官边界的关键调控因子,通过其EAR基序直接招募共抑制子TPL,形成转录抑制复合物,精确限制B类花器官基因的表达空间,确保雄蕊和心皮在正确位置发育。该发现不仅解决了长期悬而未决的SUP作用机制问题,更深化了对植物发育中转录抑制复合物工作原理的理解。值得注意的是,SUP的EAR基序在多种植物中高度保守,这一机制很可能在开花植物中具有普遍意义。此外,研究展示的精细基因组编辑策略为解析其他关键调控元件的功能提供了范本。
 生物通微信公众号
生物通微信公众号
 生物通新浪微博
生物通新浪微博
今日动态 |
人才市场 |
新技术专栏 |
中国科学人 |
云展台 |
BioHot |
云讲堂直播 |
会展中心 |
特价专栏 |
技术快讯 |
免费试用
版权所有 生物通
Copyright© eBiotrade.com, All Rights Reserved
联系信箱:
粤ICP备09063491号