综述:通过CRISPR/Cas9介导的基因编辑解锁作物对环境胁迫的韧性
《Discover Plants》:Unlocking crop resilience through CRISPR Cas9 mediated gene editing against environmental stressors
【字体:
大
中
小
】
时间:2025年11月16日
来源:Discover Plants
编辑推荐:
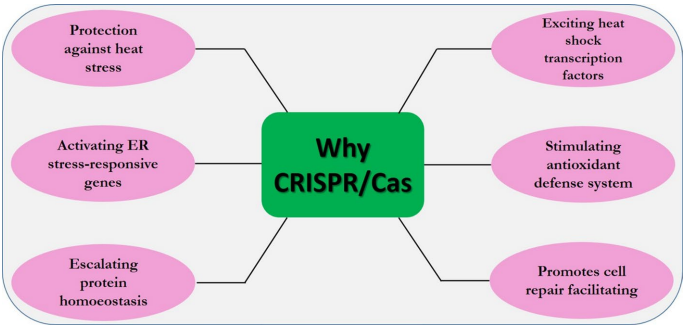
本综述系统阐述了CRISPR/Cas9基因组编辑技术在提升作物非生物胁迫(如干旱、盐碱、极端温度等)抗性方面的最新进展。文章指出,与传统育种和转基因技术相比,CRISPR/Cas9具有精准、高效、无需引入外源DNA等优势,可通过敲除胁迫负调控因子、激活有益基因、编辑关键转录因子(如DREB、WRKY、NAC)等方式,精细调控胁迫信号通路(如ABA、ROS、Ca2+信号),从而培育出具有多重抗逆性的优良作物品种,为应对气候变化下的粮食安全挑战提供了有力工具。
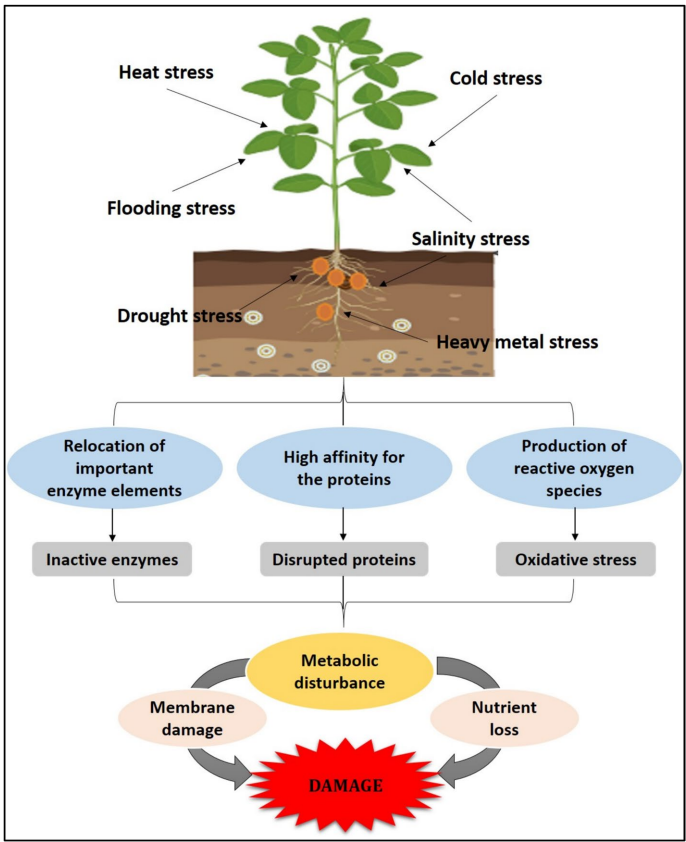
2 植物非生物胁迫
干旱、盐碱、极端温度、重金属毒性和涝害等非生物胁迫严重阻碍植物的生长、发育和生产力。这些环境限制因素会破坏生理和代谢过程,通常通过活性氧(ROS)的过量产生导致氧化胁迫,从而损害脂质、蛋白质和核酸。例如,盐胁迫导致Na+和Cl-过度积累,干扰养分吸收和酶功能,最终抑制细胞扩张和分裂。温度胁迫影响膜流动性和蛋白质稳定性――热胁迫使蛋白质变性并破坏光合作用,而冷胁迫则损害代谢活性并因冰晶形成诱导细胞脱水。
为应对非生物胁迫,植物会激活复杂的生理、生化和分子防御机制网络以恢复稳态并减轻损伤。这些机制包括形态适应(如更深的根系、叶片卷曲)、渗透保护剂(如脯氨酸、甘氨酸甜菜碱、可溶性糖)的积累以及抗氧化酶(如超氧化物歧化酶SOD、过氧化氢酶CAT、抗坏血酸过氧化物酶APX)的激活。在分子水平上,胁迫感知会触发涉及钙离子(Ca2+)、ROS、脱落酸(ABA)以及DREB、WRKY、NAC等转录因子的信号级联,这些因子调控胁迫响应基因的表达。这些基因编码脱水素、胚胎发育晚期丰富蛋白(LEA)和热激蛋白(HSP)等保护性蛋白,在胁迫条件下稳定细胞结构并维持生理功能。
3 CRISPR/Cas9概述
CRISPR/Cas9系统通过高度特异性和可编程的机制发挥作用,能够在多种生物中进行靶向基因组编辑。其核心组件是向导RNA(gRNA)和Cas9内切酶。gRNA通过碱基配对将Cas9引导至基因组中特定DNA序列,该序列附近必须有一个短的原间隔序列邻近基序(PAM),对于酿脓链球菌Cas9,PAM通常为5‘-NGG-3’。识别PAM后,Cas9解旋DNA,gRNA与互补DNA链杂交,Cas9的HNH和RuvC结构域分别切割互补链和非互补链,产生位点特异性的双链断裂(DSB)。
细胞随后通过两种主要的内源性途径修复DSB:非同性末端连接(NHEJ)或同源定向修复(HDR)。NHEJ是主要修复途径,能快速连接断裂的DNA末端,但常引入小的插入或缺失(indel),可能导致移码突变或基因敲除。HDR利用同源DNA模板进行精确修复,但效率较低。对原始系统的改进,如Cas9切口酶(nCas9)、无催化活性的Cas9(dCas9)、碱基编辑和引物编辑技术,进一步提高了编辑的特异性和精确度。
现代生物技术工具如CRISPR/Cas9允许精确编辑参与胁迫耐受性的基因,如调控干旱、盐分和冷响应的OsDREB1、NHX1和TPS1。该技术已成功应用于20多种重要农作物,能够同时靶向多个基因(多重编辑)。例如,通过编辑盐生植物中参与Na+/K+稳态、离子转运、脯氨酸积累的基因,可培育耐盐作物品种。在拟南芥中,CRISPR介导的OST2基因敲除通过提高水分利用效率增强了抗旱性。对miR169a和miR827a等microRNA的编辑也被证明可以调节耐旱途径。
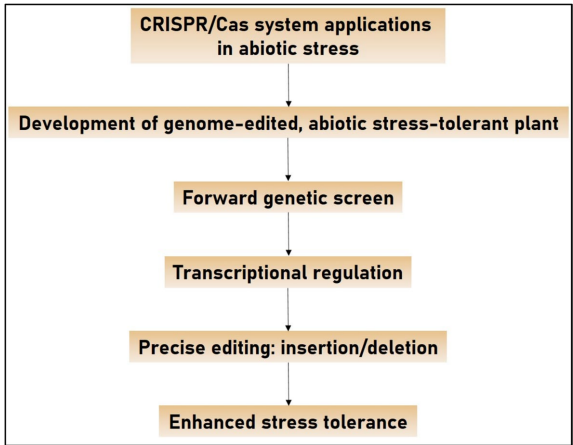
4 CRISPR/Cas与非生物胁迫
CRISPR/Cas9通过诱导靶向基因突变来增强植物对非生物胁迫的抗性。
冷胁迫与热胁迫:冷胁迫通过减少生物量、光合作用和产量严重影响植物生态。在水稻中,通过CRISPR/Cas9沉默OsPRP1会因富含脯氨酸蛋白减少而增加对冷的敏感性。冷胁迫触发CBF基因表达,进而增强COR基因以促进耐冻性。在番茄中,SlCBF1是唯一受冷诱导的CBF,其CRISPR突变体表现出更高的膜损伤,表明其保护作用。水稻中的OsAnn5和OsAnn3受顺式元件和转录因子调控,敲除这两个基因会降低耐冷性。热胁迫破坏光合作用、代谢和养分吸收。OsHSA1敲除水稻表现出热敏感性,而SlMAPK3敲低则通过HSP和HSF的表达增强耐热性。OsNAC006突变体在热胁迫下叶绿素和抗氧化活性降低。光敏色素PHYB作为热传感器,其phy突变体在拟南芥和番茄中表现出改良的耐热性。热胁迫还损害生殖,对SlAGL6、IAA9和SlDELLA的CRISPR突变可诱导单性结实,改善胁迫下的坐果。
干旱胁迫:干旱显著降低生产力。番茄中SlARF4的CRISPR敲低减少了水分流失。拟南芥AITR突变体在不影响生长的情况下增强了干旱和盐胁迫耐受性。AREBI的CRISPR激活增加了干旱韧性。玉米中ARGOS8变体改善了干旱下的产量。OsERA1、NPR1、OsPUB67和SAPK2的敲除显示出耐旱性降低,证实了它们的调控作用。这些研究表明,叠加突变可以进一步增强耐旱性。
盐胁迫:盐分由于离子毒性和渗透胁迫损害生长。CRISPR编辑的OsRR22水稻在NaCl处理下生长改善。OsVDE和DST敲除改善了ABA反应和胁迫存活。OsNAC41和SlARF4突变体显示出盐敏感性降低。AITR敲除在拟南芥和大豆中提高了耐盐性且无形态缺陷。拟南芥ACQOS突变体在盐胁迫下失去叶绿素,证实了其负调控作用。小麦中TaHAG1敲除增加了盐敏感性。马铃薯中coilin的CRISPR修饰有助于缓解盐胁迫。
重金属胁迫:镉、砷、铬等重金属对作物有毒,损害生理和分子功能。水稻中OsNramp5(锰和镉转运蛋白)的CRISPR/Cas9敲除显著降低了镉的吸收和积累。液泡重金属转运蛋白OsHMA3的编辑进一步限制了镉向地上部的转运。同样,砷解毒调节因子OsARM1的突变增强了水稻的砷耐受性。在拟南芥中,植物螯合素合成酶PCS1的CRISPR编辑降低了镉敏感性。靶向胁迫诱导型转录因子如OsNAC300和OsWRKY45也可通过调节ROS清除和金属螯合途径来增强耐受性。
5 农业中用于非生物胁迫耐受性的CRISPR/Cas9
在锌指核酸酶(ZFN)、转录激活因子样效应物核酸酶(TALEN)和CRISPR/Cas系统中,CRISPR/Cas因其简单、高效、精确而成为应用最广泛的技术。
热胁迫耐受性:环境温度升高引发细胞损伤并激活胁迫响应途径。热激蛋白、ROS清除酶和保护性代谢物的表达参与胁迫适应。例如,番茄中白化基因缺失的突变体对热更敏感。编辑TMS5基因产生了热敏感雄性不育玉米植株。BZR1的操纵通过RBOH1信号影响H2O2产生和耐热性。此外,与热敏感白化1(hsa1)基因相关的FLN2 CRISPR编辑突变体由于叶绿体发育缺陷而表现出严重的白化表型。生菜中NCED4 CRISPR/Cas9突变体在37°C下种子发芽率超过70%,耐热性显著提高。
冷胁迫抗性:冷胁迫同样有害,导致膜不稳定和氧化损伤。CRISPR技术使得能够修饰多个冷响应基因。番茄中cbf1突变体减少了电解质泄漏并保护植物免受冷害。草莓中FvICE1的过表达改善了耐冷和耐旱性,而敲除突变体则表现出抗性降低。水稻中编辑OsANN3和SAPK2基因通过ABA信号和增强膜稳定性来改善耐冷性。OsPRP1的CRISPR介导敲除导致突变体在冷胁迫下抗氧化酶活性和保护性代谢物水平降低。OsWSL5和OsZEBRA3的突变证实了这些基因在冷胁迫适应中的重要性。黄瓜中的CsWRKY46基因通过ABA依赖性途径调控冷响应基因如COR47和RD29A。
干旱胁迫缓解:干旱胁迫损害植物发育,降低产量,限制水分吸收。CRISPR在识别和修饰干旱响应基因方面发挥了重要作用。小麦中编辑TaDREB2和TaERF3基因改善了乙烯信号和抗旱性。相反,水稻中SAPK2敲除对干旱和氧化胁迫更敏感。OsERF109、OsBIERF1、OsBIERF3和OsBIERF4改变的CRISPR编辑水稻在干旱条件下生长改善。拟南芥中,AREB1、ABF1和ABF3等ABA响应基因有助于干旱适应。AREB1的过表达比敲除突变体增强了抗旱性。缺乏多个GmAITR基因的大豆突变体对干旱和盐分的耐受性更强。番茄中CRISPR介导的SIBD40和SIMAPK3沉默通过调控胁迫响应基因改善了耐旱性。靶向ARGOS8(抑制乙烯反应的基因)的玉米突变体表现出增强的耐旱性。
盐胁迫耐受性:盐胁迫破坏细胞离子平衡和水分吸收,导致氧化胁迫和细胞死亡。CRISPR使得能够功能性验证负责耐盐性的关键基因。水稻中,OsBBS1、OsMIR528和OsRAV2与盐敏感性和胁迫响应相关。SIMAP3敲除的番茄通过下调SILOX、SIGST和SIDREB表现出耐盐性。耐盐机制通常依赖于适当的离子转运。SOS1(关键的Na+外排转运蛋白)已在大豆中使用CRISPR进行编辑,导致Na+积累增加和敏感性增强。SOS1-SOS2-SOS3复合物对于盐渍条件下植物的离子稳态仍然至关重要。小麦中TaHAG1的CRISPR编辑通过增强组蛋白乙酰化改善了ROS调控和耐盐性。水稻中osbhlh024的敲除抑制了MDA和H2O2的积累,从而产生更好的耐盐性。此外,OsPIL14-SLR1蛋白调控盐胁迫下幼苗的生长,其突变体系显示出更长的根和更好的耐受性。
6 CRISPR/Cas介导的作物除草剂耐受性
除草剂是用于清除与作物竞争必需资源的有害植物(杂草)的化学药剂。近年来,基因组编辑的进步,特别是CRISPR/Cas9系统,使得能够开发耐除草剂作物品种。例如,从大豆中分离出突变基因以生产耐氯磺隆品系。乙酰乳酸合酶基因ALS1和ALS2是开发耐氯磺隆作物的主要靶标。使用单链寡核苷酸作为修复模板,已成功产生耐除草剂玉米植株。
CRISPR/Cas9介导的基因组编辑促进了耐除草剂作物的开发。乙酰乳酸合酶(ALS)是支链氨基酸(如亮氨酸、异亮氨酸、缬氨酸)生物合成中的一种酶,是磺酰脲类和咪唑啉酮类除草剂的分子靶标。基因组编辑使得玉米、拟南芥、小麦、水稻、甘蔗和番茄等作物能够获得对ALS抑制性除草剂的抗性。ALS基因密码子P171的特定错义突变(P171S、P171A、P171Y、P171F)赋予了对多种除草剂的不同程度的耐受性。在水稻中,osALS基因第1882位G-to-T颠换产生的新等位基因(G628W)赋予了对咪唑烟酸和咪唑乙烟酸的抗性。番茄中通过ALS基因的脯氨酸-186突变实现了对氯磺隆的抗性。此外,使用将胞嘧啶脱氨为尿嘧啶的激活诱导胞苷脱氨酶(AID)在水稻ALS基因中引入C287T点突变,从而产生对甲氧咪草烟的抗性。水稻EPSPS基因中的靶向基因插入和点突变也产生了耐草甘膦品种。这些CRISPR衍生的突变体在不影响产量的情况下表现出除草剂抗性。PPO1、HPPD、CP12和Ubiquitin2等基因的表达增加,表明它们在赋予水稻除草剂抗性中具有额外作用。在番茄Micro-TOM品种中,使用CRISPR-Cas9编辑除草剂靶标基因EPSPS和ALS。所得突变体的诱变效率为2-16%,并对除草剂灭草松具有抗性。CRISPR介导的腺嘌呤碱基编辑器(ABE)用于精确编辑水稻中的OsTubA2基因,产生了一个点突变,赋予了对二硝基苯胺除草剂的抗性。
7 结论
植物提供多种产品。为了提高作物生产力,植物育种者需要一系列技术。CRISPR系统比传统育种更准确、更有效。CRISPR/Cas9基因组编辑是一项真正新颖的方法。作物可以通过基因组编辑成功整合一系列遗传特征。基因组编辑技术开创了植物遗传学增强的新时代,克服了现有转基因方法的缺点。预计CRISPR/Cas9技术将有助于解决转基因生物(GMO)和社会分歧问题。
 生物通微信公众号
生物通微信公众号
 生物通新浪微博
生物通新浪微博
今日动态 |
人才市场 |
新技术专栏 |
中国科学人 |
云展台 |
BioHot |
云讲堂直播 |
会展中心 |
特价专栏 |
技术快讯 |
免费试用
版权所有 生物通
Copyright© eBiotrade.com, All Rights Reserved
联系信箱:
粤ICP备09063491号