综述:表观遗传机制调控植物对非生物胁迫的响应及其在培育气候韧性作物中的作用
《Discover Plants》:Epigenetic mechanisms regulating plant responses to abiotic stress and their role in developing climate resilient crops
【字体:
大
中
小
】
时间:2025年12月04日
来源:Discover Plants
编辑推荐:
本综述系统阐述了植物如何通过DNA甲基化、组蛋白修饰、染色质重塑及非编码RNA等表观遗传机制,动态响应干旱、盐碱、温度极端等非生物胁迫。文章重点探讨了表观遗传记忆(Stress Priming)、CRISPR/dCas9表观基因组编辑等前沿技术在作物抗逆育种中的应用潜力,并对比了印度与全球范围内的监管与伦理考量,为开发气候韧性作物提供了从分子机制到育种策略的全面视角。
植物生长常常面临干旱、高盐、极端温度等非生物胁迫的严峻挑战,这些环境压力严重制约着全球作物产量和粮食安全。有趣的是,植物并非被动承受,它们演化出了一套精妙的表观遗传调控系统,能够在不改变DNA序列的前提下,动态调整基因的表达模式,从而适应环境变化。这套系统如同一位细心的园丁,通过给DNA和组蛋白贴上不同的“化学标签”,来开启或关闭特定的基因功能。
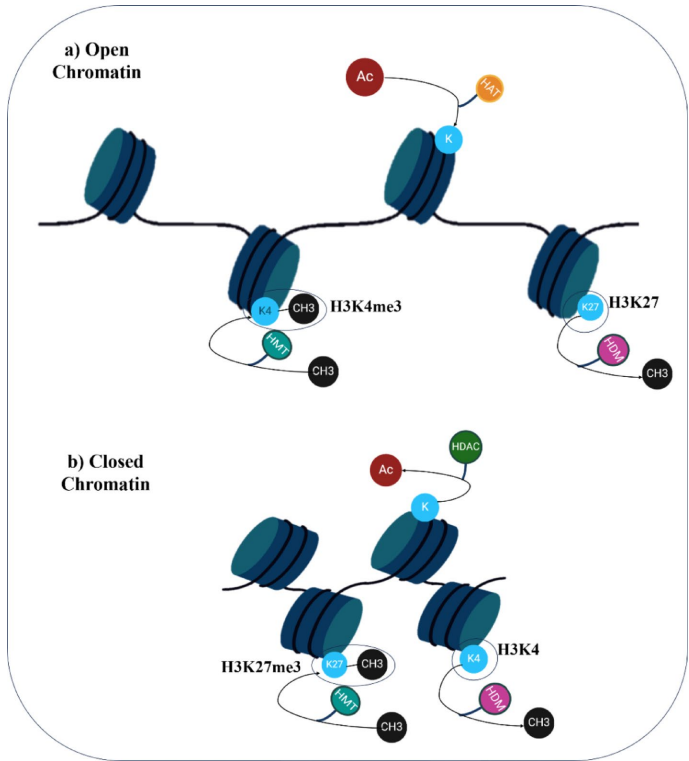
在表观遗传的世界里,DNA甲基化是其中最基础且关键的一种修饰方式。它主要指在DNA甲基转移酶(MTases)的催化下,将甲基基团添加到胞嘧啶(C)的第五位碳原子上,形成5-甲基胞嘧啶(5mC)。在拟南芥等植物中,这种修饰主要发生在CG、CHG和CHH(H代表A、T或C)三种序列背景下。其中,CG甲基化由MET1维持,通常与基因体的中度表达相关;CHG甲基化由CMT3催化,而CHH这种不对称甲基化则由DRM1/2通过RNA指导的DNA甲基化(RdDM)途径负责从头建立。当胁迫来临时,植物会灵活地运用这套系统:一方面,通过启动DNA甲基化(例如与H3K9me2相关的甲基化)来沉默那些非必需基因和跳跃基因,以维持基因组的稳定性;另一方面,又通过DEMETER(DME)和ROS1等去甲基化酶进行主动去甲基化,重新激活那些对胁迫响应和恢复至关重要的基因。更为神奇的是,一些特定的甲基化模式还能形成“胁迫记忆”,使植物在再次遭遇相似胁迫时能做出更迅速的反应。例如,拟南芥在遭受冷胁迫后,冷响应基因COR15A会呈现出持续的低甲基化状态,这使得该基因在再次遇冷时能被快速激活。
除了DNA甲基化,组蛋白修饰是另一层重要的表观遗传调控。组蛋白是DNA缠绕的骨架,其尾巴上的化学修饰能直接改变染色质的紧密程度,从而影响基因的转录活性。这些修饰包括组蛋白乙酰化、甲基化、磷酸化和泛素化等,分别由不同的“书写器”(Writers)、“擦除器”(Erasers)和“阅读器”(Readers)蛋白负责。例如,组蛋白乙酰转移酶(HATs)如GCN5,负责在组蛋白上添加乙酰基团,中和其正电荷,使染色质结构变得松散,形成开放的常染色质,促进基因转录(如H3K9ac和H3K27ac标记)。反之,组蛋白去乙酰化酶(HDACs)则移除乙酰基团,使染色质压缩,导致基因沉默。组蛋白甲基化则更为复杂,其效应取决于甲基化的位点和数量:H3K4me3通常与基因激活相关,而H3K27me3和H3K9me2则与基因 repression 和异染色质形成有关。在胁迫响应中,这些修饰动态变化:冷胁迫会减少COR15A等基因上的H3K27me3抑制性标记,便于其快速激活;热胁迫则会导致水稻中H3K9me2水平下降,并促进拟南芥中的组蛋白乙酰化,这些都与植物的耐热性增强相关。
染色质重塑复合物则是利用ATP水解产生的能量,直接推动核小体的位置或结构发生改变,如同移动书架来暴露或隐藏特定的书籍。主要的ATP依赖的染色质重塑复合物包括SWI/SNF、ISWI、CHD和INO80家族。例如,SWI/SNF复合物含有BRAHMA(BRM)和SWI3等亚基,在胁迫条件下通过核小体滑动来促进转录激活;而INO80复合物则在胁迫期间的DNA修复中发挥作用。在病原体防御中,拟南芥在受到Pseudomonas syringae感染后,会激活SWI/SNF复合物,通过重新定位核小体来促进病原相关基因的表达。在非生物胁迫响应中,染色质重塑使得胁迫耐受通路得以激活。在干旱胁迫下,SWI/SNF复合物通过滑动核小体来调控ABA响应基因如RD29A和RAB18的表达;而DDM1则保护异染色质结构以确保基因组完整性。在热胁迫响应中,BRM介导的重塑促进了热休克蛋白的表达,使转录因子能结合到热休克元件上。
非编码RNA(ncRNAs)则扮演着“分子调控师”的角色,它们本身不编码蛋白质,却能精细调控其他基因的表达。主要包括微小RNA(miRNAs)、小干扰RNA(siRNAs)和长链非编码RNA(lncRNAs)。miRNAs(如miR398)通常通过引导其靶标mRNA的降解或翻译抑制来负调控基因表达,例如miR398靶向铜/锌超氧化物歧化酶(CSD1和CSD2)以调节氧化胁迫响应。siRNAs则主要通过RNA指导的DNA甲基化(RdDM)途径,参与沉默跳跃基因以维持基因组稳定性。lncRNAs(如拟南芥中的COOLAIR和COLDAIR)则能通过招募染色质修饰酶(如PRC2)到特定基因组位点,从而改变组蛋白修饰(如H3K27me3)和DNA甲基化状态,或作为竞争性内源RNA(ceRNA)吸附miRNA,间接调控基因表达。例如,在低温胁迫下,COLDAIR的表达上调,会招募PRC2到开花位点C(FLC)基因位点沉积抑制性标记,从而调控春化作用后的开花时间。
随着高通量测序技术(如全基因组亚硫酸氢盐测序WGBS)、染色质免疫共沉淀(ChIP)和ATAC-seq等技术的发展,科学家们能够以前所未有的精度绘制植物在胁迫下的全基因组表观遗传图谱。特别是CRISPR-dCas9表观基因组编辑工具的兴起,为精准调控植物的胁迫响应提供了强大手段。与切割DNA的CRISPR-Cas9不同,催化失活的dCas9就像一个“分子导航车”,能携带各种效应结构域(如乙酰转移酶、甲基转移酶或转录激活/抑制结构域)精准定位到特定的基因启动子或增强子区域,从而在不改变DNA序列的情况下激活或抑制目标基因的表达。例如,在拟南芥中,利用dCas9与组蛋白乙酰转移酶AtHAT1融合,靶向干旱响应基因AREB1/ABF2,成功增强了植物的耐旱性;利用dCas9-VP16-p65-HSF1系统激活AVP1基因,改善了植物的干旱响应和离子积累。在水稻中,利用dCas9-TV系统对OsER1基因进行激活,实现了高达62倍的转录激活。这些案例展示了表观基因组编辑在作物抗逆育种中的巨大潜力。
在作物改良策略上,表观遗传学提供了新思路。一是利用“胁迫 priming”技术,即让植物预先经历温和的胁迫,诱导产生有益的表观遗传修饰,形成“胁迫记忆”,从而使其在面对更严重的胁迫时表现出更强的抵抗力。这种记忆有时甚至能稳定地传递给后代,即跨代遗传。二是筛选具有优良表观遗传变异的种质资源。例如,水稻的‘Pokkali’品种因其在盐胁迫下能快速调整DNA甲基化水平而表现出显著的耐盐性;通过多代干旱胁迫选育的水稻品系,也发现了与抗旱性相关的稳定表观遗传突变。这些自然存在的或胁迫诱导的表观遗传变异,可以作为表观遗传标记,用于分子标记辅助育种,加速气候韧性作物的选育进程。
将表观遗传学知识应用于作物改良,特别是在印度等面临严峻气候挑战的地区,前景广阔。印度超过50%的可耕地易受干旱、高温和盐渍化威胁,而许多主要作物(如水稻、小麦、花生、鹰嘴豆等)已显示出胁迫响应的表观遗传可塑性。印度2022年发布的生物技术指南,对不引入外源DNA的位点定向核酸酶-1和-2(SDN-1和SDN-2)编辑(包括表观遗传编辑)给予了监管豁免,为相关技术应用提供了政策空间。然而,其发展也面临挑战,包括表观遗传修饰的稳定性、潜在的脱靶效应、技术普及的公平性以及相关的伦理问题。未来研究需要更深入地探索非编码RNA与其它表观遗传层级的交叉对话,在合成生物学层面设计受胁迫诱导表观遗传调控的合成启动子,并加强田间试验验证,最终将先进的分子发现转化为农民田间实实在在的、可持续的农业生产力的提升。
综上所述,表观遗传机制为植物适应环境胁迫提供了强大的分子灵活性。通过整合对DNA甲基化、组蛋白修饰、染色质重塑和非编码RNA的理解,并利用CRISPR-dCas9等前沿编辑工具,人类正站在培育新一代气候韧性作物的门槛上。这条从基础研究到育种应用的路径,需要科学家、育种家和政策制定者的共同努力,以确保技术进步能够惠及全球农业,应对日益严峻的气候变化挑战,保障未来的粮食安全。
 生物通微信公众号
生物通微信公众号
 生物通新浪微博
生物通新浪微博
今日动态 |
人才市场 |
新技术专栏 |
中国科学人 |
云展台 |
BioHot |
云讲堂直播 |
会展中心 |
特价专栏 |
技术快讯 |
免费试用
版权所有 生物通
Copyright© eBiotrade.com, All Rights Reserved
联系信箱:
粤ICP备09063491号